Вы здесь
1.4 Современные представления о нейрогенном механизме апноэ и гастроэзофагального рефлюкса с учетом взаимодействия центральной, симп. и парасимп. нервных систем органов с собственным ритмом на примере нейрогумор. регуляции и анатом. иннервации легких
Внезапное апноэ грудных детей занимает значительное место в структуре смертности детей в возрасте до года. Хотя новорожденные имеют небольшие паузы в дыхании, настоящее апноэ представляет серьезную угрозу для их жизни. Оно возникает ввиду несовершенной центральной регуляции дыхания у новорожденных, особенно недоношенных, усугубляющейся при катаральном воспалении, гематомах верхних дыхательных путей, рините. [5; 171; 177; 178; 282]
Возникновению апноэ у новорожденных с недоразвитой ЦНС может способствовать также перегревание при нарушении нормального теплового режима. Причем, у младенцев развивается, как правило, смешанный синдром сонного апноэ. В его возникновении участвуют как центральный, так и обструктивный механизмы. Причинами развития этого состояния могут быть поражение центральных хеморецепторов или их афферентных путей, разрушение нейронов продолговатого мозга, прерывание нисходящих моторных аксонов в спинном мозге [172; 173; 181; 185; 186; 189; 239; 284].
Можно выделить два варианта сонного апноэ:
- центральный вариант обструктивного апноэ в положении пациента лежа на спине, для него характерны жалобы на храп и дневную сонливость, но в случае полисомнографии отмечаются эпизоды апноэ при отсутствии респираторных усилий грудного и абдоминального отведений, что характерно для центрального апное. Механизм этого явления можно предположительно связать с подавлением респираторного усилия спадением глотки в положении лежа на спине.
- Ларингеальный вариант сонного апноэ. Встречается, когда обструкция верхних дыхательных путей во время сна развивается вследствие нарушения иннервации гортани, приводящего к ее перекрытию, а не из-за фарингеального коллапса.
Взаимоотношение верхних и нижних дыхательных путей обусловливают нейро-рефлекторные механизмы. Импульсы, возникающие при дыхании, делятся на две группы – группа системы блуждающего нерва и группа системы тройничного и обонятельного нерва. Одновременно действуя, эти группы вызывают противоположные реакции. Раздражение блуждающего нерва учащает дыхание, уменьшая его глубину, в то же время как импульсы тройничного и обонятельного нерва замедляют и углубляют дыхание. Установлено, что выраженный бронхоспазм развивается при воздействии, как на верхние, так и на нижние воздухо-проводящие пути. При этом на 50% бронхоспазм осуществлялся сенсорными рецепторами в верхних воздухо-проводящих путях [11; 25; 32; 119; 127].
Эффект изменения бронхиального сопротивления вследствие раздражения слизистой оболочки носа различными ирритантами назван ринобронхиальным рефлексом [11].
М.С. Плужников (2002) определил, что ринобронхиальный рефлекс осуществляется через волокна сенсорной порции тройничного нерва к ядру V пары в стволе мозга, далее к двигательному соматическому ядру блуждающего нерва и в составе этого нерва к переднему и заднему легочным сплетениям. Рефлексогенная зона носовой полости функционирует в сочетании с гортанью [119]. Слизистая задней стенки гортани ниже голосовых связок при механическом раздражении вызывает кашель. Аналогичная процедура со слизистой выше голосовых связок замедляет и останавливает дыхание на выдохе без кашлевого рефлекса. При механическом раздражении рецепторов гортани возможен и бронхоконстрикторный рефлекс [11; 25; 32; 67; 68; 126].
Ночной спазм гортани появляется часто из-за раздражающего воздействия соляной кислоты на голосовые связки при гастроэзофагальном рефлюксе, возникающем ночью, и вызывающем острый стридор [182; 229; 238].
В отличии от взрослых, у которых гастроэзофагальный рефлюкс, не является жизнеугрожающим состоянием, у новорожденных и грудных детей он относится к тяжелому заболеванию и проявляется рвотой съеденной пищей [182; 238]. За координацию активности различных нервов, участвующих в акте глотания ответственен центр глотания, расположенный в близи двигательных ядер черепных нервов в продолговатом мозге. В этой же области имеется центр позыва на рвоту. Вблизи, или в самой постремальной области расположен центр рвоты [25; 32; 67; 68].
Импульсы, распространяясь по афферентным путям (волокна тройничного, языкоглоточного и верхнего гортанного нервов) достигают центра глотания, от которого по афферентным путям (веточки тройничного, языкоглоточного, подъязычного и блуждающего нервов) к мышцам, участвующим в акте глотания. Центры глотания, сердечной деятельности и дыхания, расположенные в продолговатом мозге находятся в функциональном взаимодействии. Парасимпатический и симпатический нервы оказывают стимулирующее влияние на мускулатуру пищевода [32; 48; 67; 68; 81; 255; 286; 288]. Важно отметить, что у животных после ваготомии наступает паралич пищевода, но через 9-24 часа движение в нижней трети восстанавливается, а верхняя часть так и остается парализованной [132; 139; 228; 233].
Мышечная ткань пищевода иннервируется в основном парасимпатическими волокнами блуждающего нерва в составе пищеводного сплетения. Поэтому проглатывание пищи является частично непроизвольным рефлекторным актом, который запускается при стимуляции афферентных окончаний, располагающихся преимущественно в задней стенке глотки.
Глотание начинается произвольно (ротовая полость), а с глотки и далее акт глотания непроизвольный и связан с парасимпатическыми рефлексами [11; 48; 67; 68; 117; 158; 159]. Экспериментально было доказано, что пересечение дистального отдела блуждающего нерва вызывает перистальтическую волну в гладкомышечной части пищеводa. Установлено, что перистальтические движения, стимулированные растяжением пищевода, сохраняются после ваготомии [139; 228; 233].
В пищеводе определяют верхний и нижний пищеводные сфинктеры. Дисфункция верхнего сфинктера приводит к ротоглоточной дисфагии, при которой возможны аспирация или носоглоточный рефлюкс заглатываемой пищей. Мышцы верхнего сфинктера имеют один вид иннервации – двигательные волокна блуждающего нерва [67; 68; 121; 255]
Нижний пищеводный сфинктер является условным понятием, т.к. мышца, выполняющая роль сфинктера, не обнаружена. Работы D. Liebermann-Meffert et al. (1979) позволили говорить, что сфинктерная функция связана с мышечным строением кардиального отдела желудка, в этой области имеется косое желудочно-пищеводное кольцо, обусловленное утолщением внутреннего мышечного слоя. Нижний сфинктер имеет один вид иннервации – неадренергетическую тормозящую системы (Mukhopadhyay A. K., Weisbrodt N. W., 1975).
Согласно анатомической номенклатуре и современным представлениям иннервация органов с собственным ритмом осуществляется автономной нервной системой, которая подразделяется на симпатическую (торако-люмбальную), парасимпатическую (кранио-сакральную) и метасимпатическую части, активно взаимодействующие друг с другом. Основная роль парасимпатической и метасимпатической систем состоит в осуществлении механизмов, обеспечивающих гомеостаз – относительное динамическое постоянство внутренней среды и устойчивость основных физиологических функций (Ноздрачев А. Д., Баженов Ю. И., Баранникова И. А., 2001). Депрессорный комплекс парасимпатической системы всегда вторичен.
По современным представлениям метасимпатическая система (легких в частности) – это независимая самостоятельная интегральная система, являющаяся базовой для органов, обладающих собственным моторным ритмом. Она не имеет своего центрального аппарата и ее афферентные и эфферентные связи с центральными структурами опосредованы нейронами симпатической и парасимпатической нервных систем, образующих синаптические контакты на телах и отростках метасимпатических интер- и эффекторных нейронов. Совокупность нейронов представляет собой низшие рефлекторные центры, состоящие из объединенных в функциональные модули четырех основных групп клеток: формирующие ритмическую активность (генератор ритма), эффекторные (возбуждающие и тормозящие функции нижних дыхательных путей), сенсорные. Модули объединяются тормозящими и возбуждающими связями (рис. 3, 4). В сфере управления функционального модуля находится механика гладкой мышцы, функции секретирующего и всасывающего эпителия, местного капиллярного кровотока, местных эндокринных элементов и иммунных структур [25; 32; 67; 68].
Ответственность за отклонение в гомеостазисе СО2 лежит, в основном, на центральных хеморецепторах, которые реагируют на изменения кислотно-щелочного равновесия в интерстициальной жидкости. Мониторинг парциального давления кислорода (РО2) является, исключительно, функцией каротидных телец, они получают иннервацию от каротидного синуса (Х-ая пара), узловатого ганглия (1Х-ая пара) и верхнего шейного симпатического узла [67; 68; 196].
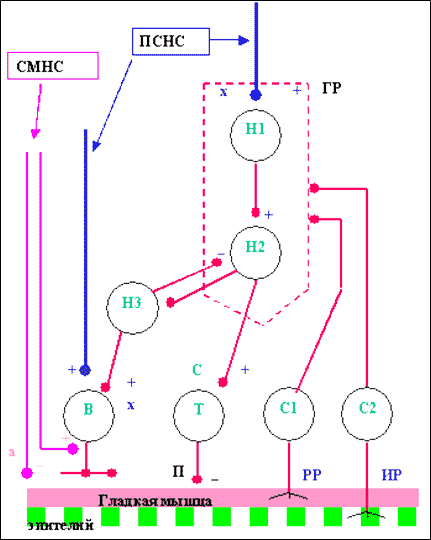
Рис. 3 . Схема функционального дыхательного модуля по Ноздрачеву А.Д. 2001.
СМНС симпатическая нервная система;
ПСНС - парасимпатическая;
ГР – генератор ритма;
Н 1-2 - нейроны генератора ритма;
Н 3 – интернейрон;
В – возбуждающий эффекторный нейрон;
Т - тормозящий эффекторный нейрон;
С 1-2 - сенсорные клетки; (+) возбуждающая передача; (-) тормозная передача;
х-холинергические,
а-адренергические,
п-пептидергические,
с-серотонергические;
РР - механорнцепторы;
ИР – ирритантные рецепторы.
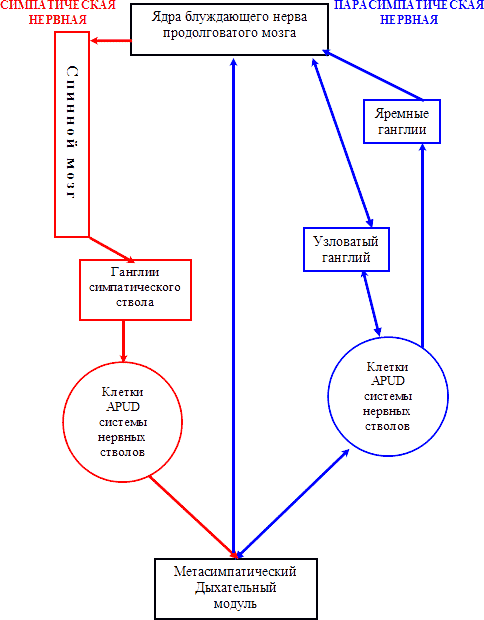
Рис.4. Схема центральной регуляции дыхательного модуля.
В системе гуморальной регуляции легких участвуют параганглии подключичной, аортопульмональной групп и мелкие скопления хромаффинных клеток в ганглиях симпатического ствола, парасимпатических узлах, в блуждающем нерве, в сплетениях грудной полости.
Все параганглии являются частью симпато-адреналовой системы. При двустороннем удалении каротидного тельца у человека полностью исчезает вентиляторный ответ.
Обширная сеть дыхательных чувствительных афферентных структур снабжает информацией дыхательный центр продолговатого мозга. Установлено, что пересечение нервной системы по понто-медуллярной границе не устраняет спонтанного дыхания, в отличии от медулло-спинальной линии. Идентифицированы три группы клеток, чьи паттерны возбуждения тесно связаны с дыхательным циклом [25; 68].
Первая группа располагается в ядре Келлихера-Фузе в клювовидно-латеральной части Варолиева моста, которая, возможно, играет роль в приспособлении дыхательного ритма к физиологической и поведенческой активности. Вторая группа локализуется в латеральном отделе ядра солитарного тракта на дорзальной поверхности продолговатого мозга и состоит из нейронов, возбуждающихся во время вдоха. Третья группа находится в вентролатеральной части, вблизи двойного ядра и состоит исключительно из экспираторных нейронов (Гриппи М.А., 2000; Ноздрачев А. Д., Баженов Ю. И., Баранникова И. А., 2001).
Таким образом, нейроны продолговатого мозга взаимодействуют с легкими посредством системы блуждающих нервов и по спино-талямическим путям со вставочными нейронами боковых рогов спинного мозга, из которых выделяются нейроны 4-го шейного сегмента – «спинальный дыхательный центр». Симпатическая и парасимпатическая нервные системы замыкаются на функциональные метасимпатические модули, в состав которых входят механорецепторы, с окончанием в гладких мышцах [25; 68].
Ирритантные рецепторы имеют чувствительные окончания между клетками эпителия, реагируют на резкие изменения объема, различные медиаторы и химические раздражители. Юкстакапиллярные рецепторы – находятся в прилегающем к капиллярам интерстиции и стимулируются интерстициальным отеком и биологически активными веществами (в том числе и нейропептидами); последние рецепторы дают начало С-волокнам, идущим, как и волокна механорецепторов, в составе блуждающего нерва. [25; 68; 93].
Из представленных литературных данных становится ясным, что по настоящее время биомеханизм и морфологические маркеры родовой травмы нервной системы у новорожденных не определены. Критерии судебно-медицинской дифференциальной диагностики родовой травмы центральной и парасимпатической нервных систем от травм насильственного происхождения отсутствуют. Морфология парасимпатической нервной системы при СВСД остается практически не изученной. В настоящее время даже при наличии самых разнообразных предположений патогенеза, ни одна гипотеза в отдельности не может объяснить феномен СВСД. Скорее можно предположить, что внезапная смерть грудного ребенка вызвана сочетанием множества факторов, возможно, что нейрогенный механизм возникновения гастро-эзофагального рефлюкса, взаимодействия верхних и нижних дыхательных путей, апноэ и другие нарушения дыхания являются проявлениями родовой травмы шейного отдела спинного мозга или системы блуждающего нерва. Представляется, что изменения симпатического дисбаланса, вследствие недостаточного или замедленного развития эфферентной активности блуждающего нерва, а также морфологические и биохимические находки при нем - глиоз и нарушение миелинизации ствола головного мозга, перивентрикулярная лейкомаляция, нарушение синтеза нейромедиаторов и гормоноподобных пептидов могут быть обусловлены родовой сочетанной черепно-спинальной травмой и, иметь соответствующие морфологические маркеры.
Читать далее раздел "2.1.Характеристика материала исследования"⇒
